124 Schistosomiasis
Schistosomiasis (bilharzia), known also by many local names such as “red-water fever,” “snail fever,” “big-belly,” and “Katayama disease,” is an “immunologic disease” induced by eggs of blood-vesselinhabiting worms of the class Trematoda, genus Schistosoma.
These eggs induce an immunologic response after they become trapped in the body organs, especially the liver, gut wall, and urogenital tract.There are three major human schistosome species: Schistosoma haematobium, which inhabit the veins of the bladder area and whose eggs are discharged in the urine; and Schistosoma mansoni and Schistosoma japonicum, which inhabit the mesenteric veins supplying the intestines and whose eggs are discharged in the feces. In every case, however, the worms may also be found in the liver and portal system. There are also a few other species that can parasitize humans. These include the japonicum- Iike Schistosoma mekongi from the lower Mekong River basin, and some African schistosomes, such as Schistosoma intercalatum, that normally parasitize cattle and wild animals.
Terminology
The terminology of this disease is very confusing. Theodor Bilharz, the discoverer of the trematode worm responsible for the disease, placed it in the genus Distoma, a broad genus that was soon abandoned as more trematode species were discovered. Numerous generic names were thereafter invented to label the worm, including Schistosoma in 1858 (the name that must stand according to the rules of zoological nomenclature), Gynaecophorus in 1858, Bilhar-
zia in 1859, and Thecosoma in 1860. Before World War II, however, in an understandable desire to honor the name of Bilharz, the disease was commonly called bilharziasis. But in 1949, members of the World Health Organization’s Study Group on Bilharziasis in Africa, ignorant of the tight rules of zoological nomenclature, recommended that the name Bilharzia be used for the worm and “bilharziasis” for the disease.
A recommendation to this effect was made to the International Commission of Zoological Nomenclature which ruled, in 1954, that the disease should be called bilharziasis, while retaining the name Schistosoma for the worms.This ruling makes little sense. The name bilharziasis would be used only for the disease if the generic name of the worm were Bilharzia, just as trypanosomiasis is used to denote the disease caused by protozoans of the genus Trypanosoma. But because the generic name Schistosoma must be retained, the term “bilharziasis” becomes invalid, although the word “bilharzia” could be used to denote the disease.
Logic, however, has frequently ruled over historical awareness, and since World War II, the Americans, in particular, have used the word “schistosomiasis,” or even its ugly derivative, “schisto,” to denote a disease whose causal agent belongs to the genus Schistosoma.
Distribution and Prevalence
The disease has an almost worldwide tropical distribution, but is mercifully absent from the Indian subcontinent. S. haematobium is highly endemic in the Nile Valley, and has an irregular distribution in the Middle East and North Africa (Map VIII. 124.1). It occurs in most West and Central African countries, along the coastal countries of East Africa from Somalia to Natal, and in the islands off the east coast. S. mansoni is highly endemic in the Nile Delta and now seems to be spreading into the Nile Valley (Map VIIL 124.2). In Africa it has a distribution similar to (although more irregular than) that of S. haematobium. But, unlike S. haematobium, S. mansoni also occurs in South America (Brazil, Surinam, and Venezuela) and in some islands in the Caribbean (Dominican Republic, Puerto Rico, St. Lucia, and others) (Map VIIL 124.3); the parasite was transported from Africa to the New World as an unintended by-product of the slave trade. Oriental schistosomiasis, caused by S. japonicum, is endemic to the Yangtze Valley and many coastal provinces of mainland China (Map VΠI.
124.4). It also occurs in Central Sulawesi and the Philippines, and there are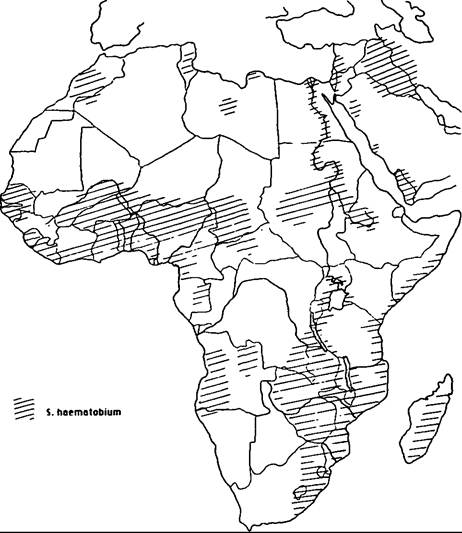
Map VIII.124.1. Distribution of Schistosoma haematobium in Africa and the Middle East. (From Farley 1991.)
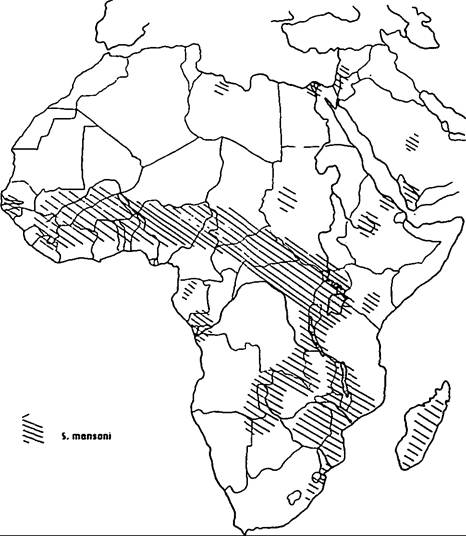
Map VIΠ. 124.2. Distribution of Schistosoma mansoni in
Africa and the Middle East. (From Farley 1991.)
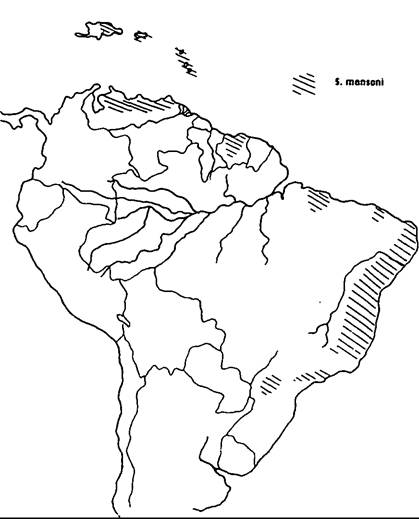
Map VIII.124.3. Distribution of Schistosoma mansoni in the Western Hemisphere. (From Farley 1991.)
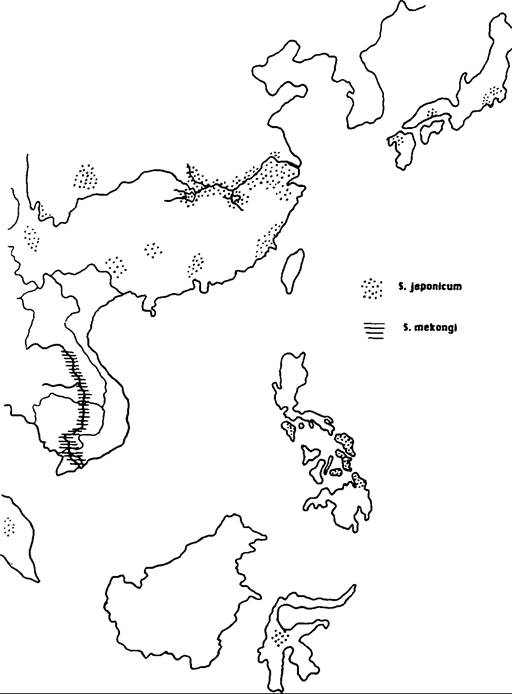
Map VIII.124.4. Distribution of Schistosoma japonicum and Schistosoma mekongi. (From Farley 1991.)
a series of smaller foci in Malaysia, Thailand, and Japan.
Over 200 million people are said to be infected with the disease, although the data on which such figures are based are extremely variable and unreliable. There is little doubt, however, that in villages of the Nile Delta and other areas where there is constant contact of the human host with water, the prevalence can be as high as almost 100 percent. On a worldwide scale, a recent World Health Organization survey revealed that in 42 countries surveyed, 21 percent of the population was infected.
Etiology
The worms are acquired in fresh water by contact with their larval stages. The worm eggs, which are highly diagnostic for each species, are shed in the urine or feces of the human host and hatch to produce a minute short-lived larval stage called a miracidium. The miracidium of each species invades the tissues of a specific snail host where it undergoes asexual reproduction, eventually to produce the final larval stage. The final stage, the cercaria, is released daily in very large numbers from the snail, swims freely in the water, and then bores into the skin of the human host.
In the human the parasite migrates to the liver via the heart and lungs, eventually to mature in the veins of the liver, gut, or bladder. Eggs appear in the urine or feces approximately 30 to 40 days after infection.Epidemiology
The disease has a very complex epidemiology, resulting in part from a quite intricate relationship between the parasite and the snail intermediate host. Not only are there strains of each schistosome species, but also there are a multiplicity of snails that vary in their susceptibility to these strains and have a taxonomy that is in a constant state of revision. Historically this has long been a source of total confusion.
In very general terms, the snail hosts of S. haematobium belong to the genus Bulinus, which has been divided into two subgenera: one with three species complexes, including the Bulinus truncatus-tropicus group; and the other with one species complex, the Bulinus africanus group. The B. truncatus-tropicus species complex act as intermediate hosts to S. haematobium in Egypt and other countries north of the Sahara, whereas the B. africanus species complex generally serve as the most important intermediate hosts south of the Sahara. The genus Biomphalaria acts as the intermediate host for S. mansoni. As in
Bulinus, the genus has been divided into species complexes, with the Biomphalaria pfeifferi group acting as the most important complex in Africa south of the Sahara and in the Middle East, and Biomphalaria alexandrina the host in the Nile Delta. In the Americas, Biomphalaria glabrata acts as the most effective host. Whereas S. haematobium and S. mansoni are transmitted by species of pulmonate freshwater snails, S. japonicum is transmitted by amphibious prosobranch snails belonging to the species Oncome- Iania hupensis, of which there are six geographic subspecies. The aquatic prosobranch snail, Tricula aperta, endemic to the Mekong River, acts as the intermediate host of S. mekongi.
Schistosomiasis can be a serious chronic disease in poor rural areas, where children and adults, because of recreational, domestic, religious, and occupational reasons, come regularly into contact with fresh water contaminated with the schistosome cercariae.
In most endemic areas, the prevalence of the disease and the intensity of infection (i.e., the number of eggs released, which is an indication of the number of worms carried) peak among those in their teenage years. Indeed in areas of high endemicity virtually all children become infected at some time. Thereafter, a decrease in prevalence and a decline in intensity occur. Even in highly endemic areas the rate of transmission of the disease is low. This follows, in part, from the patchy distribution of the surprisingly low numbers of infected snails.Reservoir hosts play an important role in Oriental schistosomiasis, where the parasite is naturally transmitted between humans and other vertebrates, including many domesticated animals such as cattle, pigs, and dogs. Although animals are believed to play little if any role in the transmission of the two schistosome species in Africa, rodents are thought to act as important reservoirs of S. mansoni in South America.
The human schistosomes belong to a large family of trematodes, the Schistosomatidae, which also parasitize birds and mammals. In many parts of the world, particularly in the lake country of the central and western United States and Canada, the cercariae of these nonhuman schistosomes can penetrate the skin ofhumans by accident. Although these cercariae are destroyed in the skin, they nevertheless cause a harmless but very irritating rash - “swimmer’s itch” or “schistosome dermatitis.”
Pathology
Disease pathology is due to embolized eggs that induce inflammatory reactions in various body organs, from which arise the classic symptoms of chronic schistosomiasis. This pathology is, however, very variable and is generally related to the intensity of infection. There are also pathological differences among the various species and among strains of the same species.
In S. haematobium lesions occur in the bladder and ureter around the entrapped and calcifying eggs, with the eventual laying down of fibrous connective tissue.
The symptoms include blood in the urine (hematuria), painful and excessive urination (dysuria), and various symptoms associated with obstructions of passages, such as distension of the ureters (hydroureter) and distension and atrophy of the kidneys through blockage of the urethras (hydronephrosis). In the intestinal schistosomes, the lesions occur in the gut wall and liver, leading to the deposition of fibrous connective tissue. The venous obstruction produced by these lesions results in a compensatory increased arterial flow that leads to portal hypertension and the classic enlargement of the liver and spleen (Iiepatosplenomegaly). Eggs of all three species may also become trapped in the lungs, and with S. japonicum, nervous disorders or “cerebral schistosomiasis” can also occur if egg aggregates come to rest in the brain.Immunology
The post-teenage decline in the disease has often been taken as evidence for a gradually acquired immunity, but this probably plays a less significant role in the decline than do changing patterns of water contact. However, “concomitant immunity” is known to occur in experimental animals. The adult worms induce an immune response that has no effect on the adult worms but that protects the host from reinfection by destroying many of the invading cercariae. The adult worms, themselves, are able to circumvent this immune response 2 to 3 days after boring into the skin by acquiring host antigens that become bound to the surface of the parasite; the host is no longer able to recognize the worms as foreign organisms. Whether concomitant immunity plays any significant role in humans has yet to be determined.
History
Pre-World WarI
Human schistosome worms were first described from Egypt by Theodor Bilharz in 1851, and their presence was related to disease symptoms by Wilhelm Griesinger shortly thereafter. Both men naturally assumed the worm to be a single species, Distoma haematobium (later named S. haematobium), occurring in the blood vessels of the gut or bladder. That there were two Egyptian species with different egg types was first suggested by Louis Sambon in 1907, who named the second species S. mansoni. A year later, Piraja da Silva in Brazil gave the first description of S. mansoni, but unfortunately assumed it to be another species distinct from both S. mansoni and haematobium. A long and testy controversy over the existence of two species in Egypt, with different morphologies and egg types, was finally resolved by Robert Leiper in 1915. Meanwhile, in 1905 Fujiro Katsurada had described eggs and worms from patients and cats in the Yamanashi district of Japan, and named them Schistosoma haematobium japonicum.
The life cycles of the worms long remained a mystery, although it was usually assumed that an intermediate host was involved. But Arthur Looss, the foremost authority on the disease at the turn of the century, argued in 1894 that there was no intermediate host, and that the miracidia bored directly back into human beings. This bizarre theory likewise generated a controversy that was finally resolved by experimental work in Japan. There, in 1913 Keino- suke Miyairi and Masatsuga Suzuki discovered the snail host into which the miracidia penetrated, and for the first time described the fork-tailed schistosome cercariae emerging from the snails a few weeks later.
Leiper had been sent to China at this time to uncover the mysterious life cycle of the worm, and, hearing of the Japanese success, had hurried to Japan to confirm the life cycle. In 1914 he was posted to Egypt, where he quickly resolved the life-cycle problem in that country, distinguishing the two species of schistosome worm in Egypt not only by their morphologies and egg types but also by their different snail hosts.
With these problems resolved, interest in schistosomiasis subsided a bit, particularly because the disease was thought curable by the drug antimony tartrate and preventable by the snail-killing chemical copper sulfate.
The Interwar Years
Between the two world wars, campaigns against the disease using these chemicals were carried out in Egypt, the Gezira irrigated area of the Sudan, Southern Rhodesia, and South Africa, particularly after schistosomiasis was found among the children of “poor whites” in rural Transvaal. In addition, the high prevalence of the disease among African mine laborers recruited for the South African mines generated research on the disease by members of the South African Institute for Medical Research. The interwar years also witnessed the first involvement with the disease by the International Health Division of the Rockefeller Foundation, which financed an attempted eradication campaign against hookworm and schistosomiasis in Egypt between 1929 and 1940. Moreover, in 1924 Ernest Faust and E. Meleney, two faculty members at the Rockefeller- funded Peking Union Medical College, first discovered the life cycle of S. japonicum in China.
Post-World WarII
After World War II, interest in the disease dramatically increased, especially in the British, French, and Belgian colonies in Africa. By 1950, schistosomiasis, previously considered to be unimportant outside Egypt, Sudan, South Africa, and to some extent China, was recognized as the most important tropical disease in the world, after malaria. In America this interest followed from an outbreak of the infection among American troops in Leyte, in the Philippines, where in 1944 over 1,000 troops of combat engineering companies involved in bridge building and road construction came down with the disease. The scientists and physicians, who were posted to Leyte to deal with the problem, brought the disease to the attention of American academics immediately after the war and initiated the growth in schistosomiasis research.
In British Africa, where medical work had long been hampered by the policies of a penny-pinching empire, a series of Colonial Development and Welfare Acts passed every 5 years between 1940 and 1955 transformed the Colonial Office into an agent of colonial development. Money was made available for medical research, most of which was allocated for work on trypanosomiasis and the tsetse fly. But a helminth subcommittee of the Medical Research Advisory Committee was also formed, which began to stress the danger of schistosomiasis, particularly as new irrigation schemes, built as part of the new emphasis on colonial agricultural development, threatened to spread the disease. The subcommittee initiated, for example, testing of a new drug, Miracil D, discovered by the British after occupation of the Bayer laboratories in Elberfeld toward the end of World War II.
The first major British work on the epidemiology of the disease in colonial Africa took place between 1955 and 1965, a period that saw the East Africa Medical Survey conducted while the Nationalist Gov- eminent of South Africa initiated surveys of the disease and supported laboratory research on experimental schistosomiasis in animals.
The 1950s also witnessed the first mass campaign against the disease that involved more than the usual introduction of latrines, drugs, and mollusc-killing chemicals by foreign experts. In 1958, as part of the Great Leap Forward, the Schistosomiasis Subcommittee of the Chinese Communist Party initiated another such campaign against schistosomiasis. Utilizing a host of methods including the mass reclamation of swamp land, the Chinese eradicated the disease from many areas of the country. In 1949 perhaps 10 million Chinese were heavily infected with S.japoni- cuτn, whereas today the number is reported to have dropped to 2.4 million lightly infected cases.
The threat posed by irrigation schemes led to the postwar development of better molluscicides to replace the ineffective copper sulfate. Niclosamide, selling under the trade name OfBayluscide1 became the chemical of choice, but results were disappointing. Chemotherapy has now become the favored weapon, particularly after the important 15-year experimental control campaign in St. Lucia (1966-81), sponsored by the Rockefeller Foundation, and after the development of single-dose drugs to replace the highly toxic antimony compounds and Miracil-D, used earlier. Oral Oxamniquine proved effective against S. mansoni, and metrifonate against S. haematobium, whereas the newly developed praziquantel is emerging as a “wonder drug” effective against all schistosome species.
Today, with major support from the McConnnell Clark Foundation, a great deal of emphasis is being placed on problems of schistosome immunity and the development of vaccines.
John Farley
Hoffmann, D. B., and K. S. Warren. 1978. Schistosomiasis TV. Condensation of the selected literature, 1963—75. Washington, D.C.
Jordan, Peter. 1985. Schistosomiasis: The St. Lucia project. Cambridge.
Jordan, Peter, and Gerald Webbe. 1982. Schistosomiasis: Epidemiology, treatment and control. London.
Kean, B., et al., eds. 1978. Tropical medicine and parasitology. Classic investigations. Ithaca, N.Y.
Maldonado, J. F. 1967. Schistosomiasis in America. Barcelona.
Nelson, G. 1977. A milestone on the road to the discovery of the life cycles of the human schistosomes. American Journal of Tropical Medicine Hygiene 26: 1093—100.
Report of the American schistosomiasis delegation to the People’s Republic of China. 1977. American Journal of Tropical MedicineHygiene 26: 427-57.
Sandbach, F. R. 1976. A history of schistosomiasis research and policy for its control. Medical History 20: 259-75.
1977. Farewell to the god of plague - the control of schistosomiasis in China. Social Science and Medicine 14: 27-33.
Sasa, M. 1972. A historical review of the early Japanese contributions to the knowledge of schistosomiasis japonica. In Research in filariasis and schistosomiasis, ed. M. Yokogawa, Vol. 2, 235-61. Baltimore.
Warren, K. S. 1973. Schistosomiasis. The evolution of a medical literature, selected abstracts and citations, 1852-1972. Cambridge.
Warren, K. S., and D. B. Hoffman. 1978. Schistosomiasis III: Abstracts of the complete literature, 1963—1974. Washington, D.C.
Warren, K. S., and A. Mahmoud. 1967. Tropical and geographical medicine. New York.
Warren, K. S., and V. A. Newill. 1967. Schistosomiasis: A bibliography of the world’s literature from 1852 to 1962. Cleveland, Ohio.
Bibliography
Abdel-Wahab, M. F. 1982. Schistosomiasis in Egypt. Boca Raton, Fla.
Basch, P. 1986. Schistosomiasis in China: An update. American Journal of Chinese Medicine 14: 17-25.
Cerqueira Falcao, E. de. 1953. Novas achegas ao estudo da determinaςao da especificidade do S. mansoni. Rio de Janeiro.
Farley, John. 1988. Bilharzia: A problem of native health. In Imperial medicine and indigenous societies, ed. D. Arnold, 189—207. Manchester.
1991. Bilharzia: A history of imperial tropical medicine. Cambridge.
Hartwig, G., and K. Patterson. 1984. Schistosomiasis in twentieth century Africa: Historical studies on West Africa and Sudan. Los Angeles.