Although the antiquity of the earliest colonists in the Americas remains a controversial subject, human presence is well established by the close of the Pleistocene, approximately 10,000 B.P. (before the present) (Bryan 1978, 1986; Shutler 1983; Fagan 1987).
Kill sites and habitation areas, in conjunction with lithic tools and a few human remains, provide convincing evidence for the presence of highly mobile, small groups whose subsistence was based on hunting and gathering of naturally available “wild” resources.
The health of these most ancient American populations, poorly documented owing to a paucity of human remains (Hrdlicka 1907, 1912; Stewart 1946, 1973; Young 1988), can best be inferred by analogy with recent hunting and gathering peoples. In making inferences we must keep in mind that such groups today tend to occupy marginal environments, unlike the often resource-rich ecosystems that attracted early human populations. If we use contemporary hunter-gatherers for our model, then parasitic infections, infections for which insects and animals serve as the primary vectors or intermediate hosts, and traumatic episodes would have been among the primary sources of ill health among the earliest Americans (Dunn 1968; Truswell and Hansen 1976; Howell 1979; Lee 1979; Cohen 1989). Degenerative diseases, neoplasms, and epidemic diseases would have been extremely rare, as would have been chronic undernutrition. Seasonal periods of nutritional stress would, however, have been expected. Thus, health status would have reflected the exceptionally close relationship between huntergatherers and their environment.
We must be careful, however, not to overgeneralize and thus simplify our picture of huntergatherer adaptations, which were undoubtedly complex and rich with the knowledge gained through intimate acquaintance with the landscape (Cohen 1989). Given the wide variety of settings available for human occupation in the Americas, ranging from stark Arctic polar environments to lush tropical forests, we should anticipate that human cultural and biological adaptions ranged widely within the context of changing cultural systems and environmental regimes.
The quality of life, as measured by health status, would have depended on the nature of these interactions as well as those among the human groups themselves.Beginning with the earliest colonists - the Americans who entered this continent before the end of the Pleistocene - we find a rich fabric of evolving relationships between people and their natural and cultural environments that held strong implications for both health and the history of disease. Although certain groups maintained life-styles without plant and animal domestication (or management) throughout their history, the Americas also saw the development of complex civilizations. The Maya, the Aztec, the Inca, and - on a smaller scale - the eastern North American agriculturalists, known to archeologists as Mississippian peoples, all developed cities where thousands of people lived together in permanent settlements. Problems of health, sanitation, and nutrition common to such large agglomerations of humans throughout the world emerged in each of these situations.
Although animal husbandry never assumed the importance that it did in the Old World, the dog was one of several domesticated New World animals, along with the turkey, the Muscovy duck, the guinea pig, the alpaca, and the llama (Crosby 1972). These animals, living close to humans, could have served as vectors or intermediate hosts of disease. Similarly, the nutritional adequacy of ancient diets depended on these as well as wild animal resources.
Inedible, but economically important plants, such as cotton and tobacco, along with many comestibles including maize, manioc, squash, beans, cocoa, sunflower, potatoes, chiles, chenoa, and tomatoes, were domesticated (Crosby 1972). These crops attracted the attention of Europeans and are presently part of Western cuisine (Crosby 1972, 1986). Other indigenous plants were also cultivated prehistorically, with some being truly domesticated, that is, showing distinctive morphological changes due to human intervention or growing beyond their natural range.
In eastern North America, for example, a group of cultivated indigenous plants, termed the eastern horticultural complex, included sumpweed (Iva annua), goosefoot (Chenopodium berlandieri), knotweed (Polygonum erectum), and little barley (Phalaris carolin-Figure V.8.1. Model time line for prehistoric eastern North America. (Based on Buikstra 1988.)
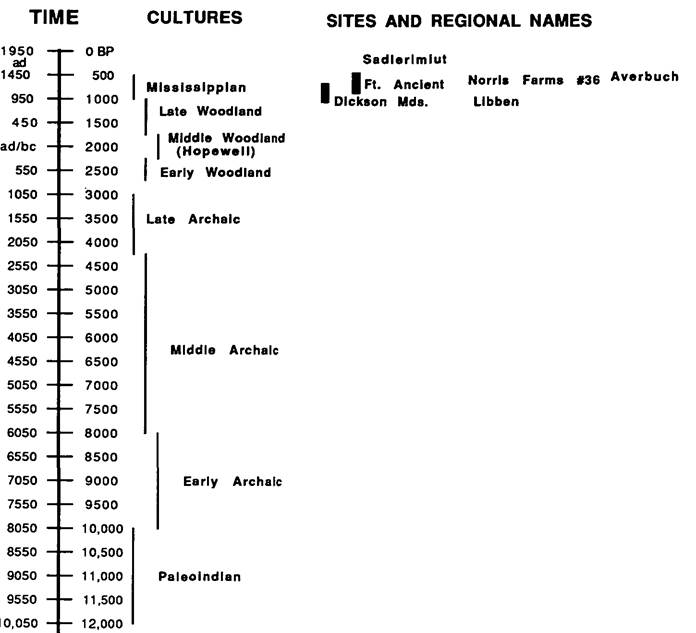
iana), as well as sunflower (Heliantus annuus) and various cucurbits (Asch and Asch 1985; Smith 1987). In that nutritional adequacy is closely linked to health status, these components of ancient diets must also be mapped when one is investigating paleo- pathological conditions.
This review of health conditions in the preColumbian Americas focuses primarily on the evidence gained through the study of skeletal material. Mummified remains, a very rich data source, are unfortunately found only in a few settings favoring soft-tissue preservation, especially the arid North American Southwest and coastal Peru and Ecuador. Dry caves, such as those of the Mammoth Cave system in North America, also yield mummified human remains, but only in small numbers (Watson 1974). Other sources of information concerning disease and medical treatments, such as the Mochica and Mexican figures, and various pictographs will not be considered here. These renderings are notoriously ambiguous, frequently charged with symbolic meanings that relate to ritual life rather than accurate representations of maladies and deformities.
Figures V. 8.1 and V. 8.2 present model chronologies for eastern North America and coastal Peru and Ecuador, two regions frequently referenced in the following text. The chronological basis for Figure V. 8.1 follows J. Buikstra (1988); Figure V. 8.2 is based on the sequences presented in R. Keatinge (1988) and B. Fagan (1972). Sites discussed in this text are placed appropriately on these time lines. This brief chronological review is developed in order to establish a broad time scale against which to display disease patterning.
In general, North American materials are emphasized, given that the majority of the relevant literature is based on North American collections.We first focus on trephination, distinctly South American and an enormously successful form of ancient surgery. Following this we consider the expression of two forms of trauma in human skeletal materials, fractures and osteoarthritis. A discussion of fractures and true degenerative joint disease centers on North American materials, as does a consideration of rheumatoid arthritis in adults. Brief mention, however, is made of an example of juvenile rheumatoid arthritis that was recovered from a c. IOOO B.P. Peruvian grave. The subsequent discussion of two forms of infectious pathology, trepone- matosis and tuberculosis, ranges widely across the hemisphere, as does the description of techniques for inferring diet from bones and teeth. We close with a consideration of parameters used to estimate population health, dealing primarily with North American

Figure V.8.2. Model time line for prehistoric Peru and
Chile. (Based on Keatinge 1988 and Fagan 1972.)
skeletal samples that chronicle the development and intensification of maize agriculture.