Allochthonous Parasites in the Americas
Previous Syntheses
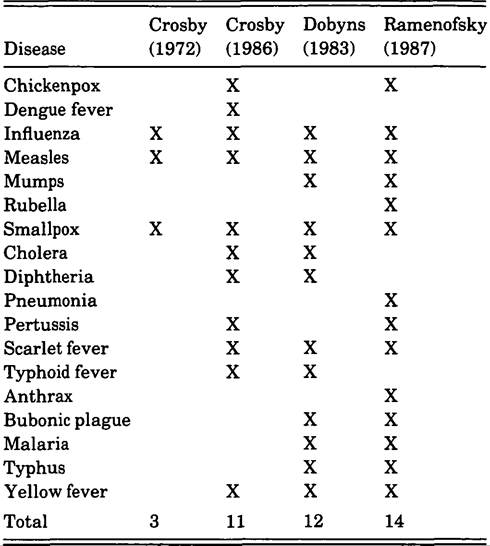
Some researchers have investigated the introduction of specific diseases to the Americas in general Table V.9.1. Diseases suggested to have been introduced to the Americas
(e.g., Stearn and Stearn 1945), whereas others have studied the introduction of all diseases to a specific region (e.g., Cook 1982).
Few have attempted a comprehensive treatment of the introduction of all diseases to both continents. Table V.9.1 summarizes the conclusions of three scholars regarding the introduction of acute infectious agents to the Americas.It will be noted that none of the lists includes epidemic (venereal) syphilis or tuberculosis. The omission of these two major diseases should not be interpreted as supporting the notion that either disease originated in the Americas. Controversies over their origins are extensive (regarding syphilis: Crosby 1972; Ramenofsky 1987; Baker and Arme- Iagos 1988; regarding tuberculosis: Black 1980; Clark et al. 1987; Ramenofsky 1987). Until this question is resolved, these diseases cannot be considered to be either allochthonous or autochthonous to the New World.
There are some disagreements about specific introductions that stem from the temporal or intellectual focus of the work. In 1972 A. Crosby considered only sixteenth-centmy introductions. In 1986 he adopted a much larger temporal framework; his suggested introductions reflect that change. Although in 1983 H. Dobyns described more introduced infections than those listed in Table V.9.1, the summary is limited to those diseases that arrived during the sixteenth and seventeenth centuries. My own list (1987) is also limited to the first two centuries of European contact.
In developing that list, I weighed evidence from all sources described previously.
Although my total was greater than either Crosby’s or Dobyns’s, several diseases were excluded; other diseases were included. Cholera was excluded because according to current opinion (Creighton 1894; Benenson 1976a) it spread to the Americas in the eighteenth century. Diphtheria was excluded because of the ambiguity of the name (Creighton 1894).Typhoid (enteric) fever was excluded for several reasons. Although native populations of the Americas certainly suffered from diarrheal infections in the postcontact period, the question becomes whether these infections were new and whether Salmonella typhi caused some of the infections. The first question cannot be answered; the second can be addressed indirectly. The disease was not isolated from typhus or other nonspecific childhood fevers until the nineteenth century (Creighton 1894; Overturf and Underman 1981). In addition, although rare, typhoid fever can take a chronic form and can spread to humans from nonhuman reservoirs including turtles (Youmans et al. 1980). The combination of a chronic state and the indisputable presence of turtles in precontact America suggested that S. typhi could have been present in the Americas before 1492.
The diseases that I listed as introductions, but not treated by Crosby or Dobyns, include anthrax, rubella, and pneumonia. Anthrax is typically a relatively minor infection in humans. Domestic stock, cattle, sheep, horses, and goats are the primary source of the infection; humans become ill through accidents of proximity (Whiteford 1979). In those areas, such as the Southwest or the pampas of South America, where domestic stock were part of the cultural baggage of European colonists, anthrax could have been a source of infection.
Rubella was added to the list for two reasons. First, although a relatively mild disease in adults, it can seriously affect the reproductive fitness of a population. If a woman develops German measles during the first trimester of pregnancy, the fetus may be bom with major congenital defects, including cataracts, heart disease, microcephaly, and mental retardation (Top 1976).
Second, although rubella was confused with measles and smallpox until the eighteenth century, the presence of measles and smallpox in America during the sixteenth and seventeenth centuries suggested that rubella could have been introduced at the same time.The bacterial pneumonias are caused by a group of unrelated organisms. Some of these (Streptococcus pneumoniae) are part of the normal flora of the human upper respiratory tract. Thus, some causative agents of severe lung infections in humans had to have been present in the Americas before European contact. In contrast, contemporary studies (Brenneman et al. 1987) suggest that some native American populations are at higher risk from other agents of pneumonia (Hemophilis influenzae). This elevated risk may indicate that not all causative bacteria of pneumonia were present before Europeans arrived in the Americas. Moreover, because pneumonias are major secondary invaders that follow such viral infections as measles and influenza, and because both measles and smallpox were allochthonous to the Americas, it is likely that at least some types of lung infection spread to the Americas with Europeans. As health deteriorated from viral infections, bacterial agents could invade and cause death.
Although both Dobyns and I view epidemic typhus as an Old World introduction to the Americas, H. Zinsser (1947) thought that it predated European contact. The vector of typhus is the human body louse. The current worldwide distribution of people means that epidemic typhus is also worldwide (Wisseman 1976). Although it is likely that body lice predated Europeans in the Americas, I think it unlikely that typhus-infected lice were present. The disease thrives under conditions of intense crowding, poor sanitation, and social or economic upheaval associated with war. Despite the archeological evidence of warfare in the Americas, the distribution of human populations was inappropriate for fixing epidemic typhus as part of the disease load of native populations.
Dense concentrations of people were simply too rare.In summary, a minimum of 11 and a maximum of 14 viral, bacterial, or protozoal diseases are suggested in Table V.9.1 as having diffused to the Americas during the first two centuries of European contact. There is unanimous agreement on five disease introductions (influenza, measles, smallpox, scarlet fever, and yellow fever). I have increased the total introductions from 11 to 14 by adding anthrax, rubella, and pneumonia.
Current Synthesis
After reexamining the evidence for and against the introduction of diseases listed in Table V.9.1,1 have removed one disease. Chickenpox is omitted from Table V.9.2 because it is present in small nucleated populations that lack antibodies for acute, infectious Table V.9.2. Viral, bacterial, and protozoal agents introduced to the Americas

microbes (Black 1980). In addition, Varicella zoster can follow a chronic course, expressing itself as chickenpox in children and as shingles in adults (Brunell 1976).
Table V.9.2 lists viral, bacterial, and protozoal agents, characterized by two modes of transmission. Eight of the 13 viruses (influenza, measles, mumps, rubella, smallpox) and bacteria (pneumonia, scarlet fever, pertussis) colonize humans only and are transmitted by them only. Of the remaining 5 diseases, yellow fever is viral; anthrax, plague, and typhus are bacterial; and malaria is protozoal. All 5 of these agents are classified as zoonoses, meaning that the primary reservoir is a nonhuman, invertebrate or vertebrate species (except in rare cases, e.g., pneumonic plague [Benenson 1976b]).
Previously discussed difficulties of reconstructing past epidemics by a single method have direct implications for the current synthesis. I will not attempt to reconstruct the precise date or port of entry of each disease introduction. My goal is to build general expectations about the postcontact spatial persistence of parasites, and to accomplish this I will use data on transmission cycles, evolution, ecology, and history.
For those viruses and bacteria that colonize and reproduce only in humans, density of human populations, regularity of communication, and incubation period of the parasite determined the size of spatial epidemic waves. As population density decreased or communication became irregular relative to the incubation period of the parasite, the parasite died out. Whether a second introduction of the same parasite caused another epidemic outbreak varied according to the number of reproductively active survivors and time. Although individuals who escaped infection during the first disease event could be subsequently infected at any time, the size of the susceptible pool determined whether any new outbreak would be local or regional in scale (Bartlett 1956; Black 1966; Cockbum 1971).
Recently, the 1520 introduction of smallpox to the Americas has been a subject of some interest. It is a classic example of disease transmission and spatial diffusion. On the basis of information drawn from historical documents, Dobyns (1983) has argued that the virus became the initial New World pandemic, spreading as far south as Chile and as far north as the Canadian border. The Caribbean islands were the initial focus of infection in 1518. The virus was then carried to Mexico by a crew member of Panfilo de Narvaez’s expedition. The large size and extreme density of aboriginal populations in the valley of Mexico encouraged the rapid dissemination of the virus.
The question of spatial diffusion beyond the valley of Mexico has been investigated by historians and archeologists. Noble Cook (1981), historian, has not found evidence of spatial diffusion into the Peruvian Empire. A second introduction of smallpox to Panama in 1524 did, however, spread down the Andean chain (Cook 1982). Using climatic data to explain the persistence of smallpox, S. Upham (1986) argued that the smallpox epidemic did spread to southwestern groups. Other archeologists (Ramenofsky 1987; Campbell 1989) have relied on archeological indicators of population change and time.
Although generally supportive of an early-sixteenth-century introduction of the parasite to North America, the archeological evidence of catastrophic population loss was ambiguous. Evidence of a sixteenth-century disease introduction was not documented in central New York or the Middle Missouri (Ramenofsky 1987). S. Campbell (1989) documented a large population decline in the Chief Joseph Reservoir during the early sixteenth century. I discovered a comparable population decline in the Lower Mississippi valley between 1540 and 1600. The magnitude and rapidity of the loss argued in favor of some acute, infectious parasite. Neither the 1520 nor the 1524 smallpox epidemic could be implicated directly.In summary, for directly transmitted viruses and bacteria, the distribution of susceptible populations, communication systems, and incubation periods of the parasite determined whether local disease events became regional or multiregional. Even with detailed historical records, it is difficult to define the spatial extent of a specific parasite. Although the scale of resolution currently obtainable from regional archeological data bases may be sufficient for concluding that acute infectious microbes caused catastrophic die-offs, current knowledge is simply insufficient for stipulating whether one or another epidemic event was causal.
Transmission cycles of zoonotic infections are fundamentally different from infections transmitted solely between humans. Because vertebrate or invertebrate species other than humans are the primary reservoir of the parasite, niche requirements of the reservoir and microbe determined the postcontact spatial pattern across the American continents. Humans became infected through accidental interactions with the reservoir and causative agent. The following focuses on those aspects of reservoir or microbial niches pertinent to reconstructing the postcontact spatial patterning of these infections.
Temperature severely Cintails the distribution of yellow fever. The viral disease is transmitted to humans or other nonhuman primates by several mosquito vectors, including Aedes and Haemagogus (Taylor 1951). The optimal temperature for incubation of the arbovirus and the transmission ability of the vectors is approximately 30oC (Whitman 1951). At temperatures less than 24°C Haemagogus, for instance, cannot transmit the disease, and the incubation period of the virus lengthens. In addition, the vectors have preferred breeding habitats. Aedes aegypti breeds in clay-bottomed containers; other species of Aedes and Haemagogus prefer breeding in trees of climax rain forests (Carter 1931).
Like yellow fever, malaria is a vectorbome disease that is transmitted to humans by numerous species of Anopheles. The temperature limitations and elevational preferences of these species vary greatly. Whereas Anopheles maculipennis can reproduce in cold pools along lake margins at elevations greater than 4,000 feet in the western United States, Anopheles quadramaculatis prefers breeding in swampy nonbrackish pools or bayous typified by the Lower Mississippi valley (Hackett 1941; Watson and Hewitt 1941). In addition, the body temperature of the vector affects the reproductive potential of the protozoans. Plasmodium υiυax and Plasmodium malariae will not develop in anophe- Iines if the body temperature is less than 15oC; Plasmodium falciparum requires an anopheline body temperature of greater than 18oC for reproduction (Zulueta 1980).
Epidemic typhus is an acute infectious disease for both vector and human populations. The human body louse, Pediculus humanus, is the vector; the causative agent is a bacterium, Rickettsia prowaze- kii. After ingesting the agent, the typhus-infected louse dies within 7 to 10 days. When the louse feeds on human blood, the rickettsial agent is transmitted to humans (Zinsser 1947; Wisseman 1976).
Plague and anthrax are bacterial infections transmitted to humans from nonhuman mammalian reservoirs. The distribution of the reservoir largely determines the distribution of the disease. Wild rodent populations in wooded or desert areas are the true reservoir of the plague bacterium: In these settings, the disease is endemic (Meyer 1963; Benenson 1976b). In urban areas, the causative agent, Yersinia pestis, is transmitted to humans through a secondary vector, the rat flea, Xenopsylla cheopis. The proximity of humans to rats and their fleas creates a tertiary focus of the disease.
As previously mentioned, anthrax is a bacterial disease of domestic ungulates. The causative agent, Bacillus anthracis, is transmitted to humans through the ingestion of contaminated meat or milk products, contact with infected animals, or the inhalation of viable airborne spores (Brachman 1976; Whiteford 1979).
Because not all New World habitats were appropriate for the reproduction of reservoirs, vectors, and parasites, it is likely that zoonotic infections were more limited spatially than were directly transmitted viruses and bacteria. Yellow fever became fixed in tropical climates; malaria survived between 60o north latitude and 40o south latitude (Watson and Hewitt 1941); epidemic typhus survived where human groups were concentrated; herds of domestic ungulates became the focus of anthrax; bubonic plague was fixed as a disease of ports. As rats migrated away from ports, plague also migrated.
The variability in reproductive requirements and, therefore, transmission cycles of parasites has implications for the postcontact spatial patterning in the Americas. Because all native populations were virgin soil, the accidental introduction of any parasite could decimate a single population aggregate. Whether the parasite diffused spatially and acquired an endemic status in the rich American environment depended on a number of factors.
The probability of fade-outs of directly transmitted viruses and bacteria varied according to stochastic contacts between infected and susceptible hosts, the overall distribution of population, and the time separating introductions of the same parasite. As just described, the spatial pattern of zoonoses depended on reproduction requirements of reservoirs, vectors, and parasites. Certainly by the seventeenth century, natural selection had fixed the spatial variability of all introduced parasites. Like threads joining patches of a quilt, communication mechanisms linked sources of infection to potential recipients. Each epidemic wave further reduced survivors of previous disease outbreaks.