Ancient DNA in Europe
Ancient DNA (aDNA) is the final piece of the phylogeographic jigsaw, and has seen a parallel revolution to that in genomics over the past decade - also driven primarily by improvements in technology.
The analysis of aDNA has the potential not only to test models built on the basis of modern variationCaption for Figure 2.3 (cont.)
subgroups of genotypes within a sample set with distinctive allele frequencies. Each individual is represented by a narrow vertical column, separated into coloured segments representing the individual's estimated membership fractions for each of K = 14 clusters. Black lines separate individuals belonging to different populations. Populations are labelled below the figure, with their broader affiliations above it. We conducted the analysis with a random seed number generator on the linkage-disequilibrium (LD) pruned data set (to remove non-independently segregating alleles) at K = 2 to 16. K = 14, which had the lowest CV score, is the one shown. All data are from Illumina 610K and 650K chips with 544,193 SNPs; after LD pruning the total number of SNPs was 234,699. and archaeology, but also to expand upon the details, as alluded to above. Moreover, it opens up questions less accessible to studies of modern variation, such as small-scale movements in later prehistory, and most excitingly of all it can reveal processes that have been erased by time, such as population extinctions.
This new phase of archaeogenetics has once again brought new voices to the debate. If the late 1990s and early 2000s were dominated by discussion of the mtDNA and Y-chromosome results from contemporary populations, and emphasized continuity between Neolithic and Mesolithic populations in Europe, in the years from 2005 onwards ancient DNA took the leading role, and the talk was now of discontinuity.
The game-changer was the work of Haak and colleagues on mtDNA control-region sequences from LBK burials in central Europe.[39] Although 2005 was also the year in which ‘next-generation' DNA sequencing technologies were launched, the work of Haak et al.
was rather on the careful application and modification of ancient DNA techniques developed over the preceding fifteen years. What was revolutionary about this work was the outcome: that a large fraction of the LBK mtDNA lineages belonged to a cluster, haplogroup N1a, which was vanishingly infrequent in the modern population. This was clearly not the result of a single rare modern contaminant - the N1a lineages represented were quite diverse. At a stroke this both answered doubts about contemporary contamination that had plagued previous ancient human DNA studies and pointed to something completely new.Although the initial interpretation invoked not discontinuity with the Mesolithic but continuity, this was revised when the researchers began to obtain sequences from Mesolithic and late Palaeolithic samples.[40] The mtDNA variation in contemporary Europeans had led to the prediction that haplogroup U, and especially U5, was likely to have been among the earliest modern human lineages in Europe, in the early upper Palaeolithic. U5b was thought to have expanded from southwestern into northeastern Europe at the end of the last Ice Age, and U4 to have expanded in the eastern European plain at about the same time.[41] The new data from Mesolithic and late Palaeolithic specimens was indeed heavily enriched for haplogroup U, especially U5 and U4, and particularly in eastern Europe, where sixteen samples from Poland, Lithuania, and Russia all belonged to U4 or U5. Mesolithic specimens from Germany also showed a high level of haplogroup U, but with haplogroups J, T, and K also present (although restricted to a single late site regarded as a Mesolithic enclave). Since the lineages in Neolithic central Europe were more diverse, this pattern suggested a sharp Mesolithic-Neolithic discontinuity, with replacement of foraging populations by new populations.
Once again, the striking differences between the Mesolithic samples and modern populations lent credibility to the results.
Although the potential for contamination remains significant, the tide seemed to have turned. Even so, despite all the precautions taken, the samples used were museum specimens, often excavated a very long time ago - a risky strategy, because the sources of potential modern human DNA are so ubiquitous that it is virtually impossible to exclude from specimens that have not been excavated in sterile conditions. The presence of identical sequence haplotypes seen in several studies, for example, indicates that we should remain cautious.Nevertheless, the significance of the results cannot be understated. Nia mtDNAs appear to specifically mark the LBK, and have in fact only been seen in one Neolithic individual outside of central Europe, in a megalithic burial chamber in western France (implying immigration into that region from the LBK zone).30 The source of the central European Neolithic lineages, however, remains controversial. Although Haak et al. argued for an Anatolian origin, modern Nia lineages in Anatolia are rare and match extant Central and South Asian lineages, whereas the LBK samples all fall within a European subclade, Niai. A founder analysis of Nia has suggested it may have spread from the Near East into Europe as early as the late Glacial, matching the best estimate for the age of the European-specific subclade of c. 17 ka.3i
These studies of control-region variation have been followed up in even more spectacular fashion by a study of thirty-nine prehistoric central European whole haplogroup H mitogenomes, from the early Neolithic to the Bronze
phylogeny in eastern and western Slavs', Molecular Biology and Evolution, 25 (2008), 1651-8.
30 M.F. Deguilloux et al., ‘News from the west: ancient DNA from a French megalithic burial chamber', American Journal of Physical Anthropology, 144 (2011), 108-18.
31 M.G. Palanichamy et al., ‘Mitochondrial haplogroup N1a phylogeography, with implication to the origin of European farmers', BMC Evolutionary Biology, 10 (2010), 304; V.
Fernandes et al., ‘The Arabian cradle: mitochondrial relicts of the first steps along the southern route out of Africa', American Journal of Human Genetics, 90 (2012), 347-55.Age.[42] Haplogroup H is both the most common modern European hap- logroup, at >40 per cent, and also one of the most poorly resolved at the level of control-region sequences, so whole-mtDNA sequencing is particularly valuable. Again, the nature of the results renders contamination very unlikely to be a significant issue with these data.
Intriguingly, the H lineages appear to become dramatically more diverse from the middle Neolithic (c. 6 ka) or late Neolithic onwards, suggesting a change in the gene pool during the course of the Neolithic that might mirror the depletion of Nιa. The early Neolithic samples cluster with modern Near Eastern populations and the late Neolithic samples with modern Iberians in PCA, leading the authors to again suggest immigration from the Near East with the early Neolithic and substantial changes thereafter from the southwest. However, such frequency-based analyses are precarious with so few data, and the presence in the early Neolithic of the Hi cluster, which is thought to have spread from Iberia in the Mesolithic,[43] points clearly to assimilation of Mesolithic lineages. Indeed, in the context of the modern distributions, many of the other early Neolithic lineages appear more likely to have a European than a Near Eastern source, with the low diversity perhaps the result of a dramatic expansion from few founders.
The case remains open, but leapfrog migration of Neolithic pioneers from Anatolia into southeastern Europe, with an archaeologically attested pause for an adaptation of farming technology accompanied by assimilation of indigenous populations (carrying, for example, Nia and H lineages), followed then by a rapid expansion into central Europe as the LBK, might well be the best explanation for this pattern.[44] The pioneers would have imported cattle[45] and pigs[46] from the Near East, although the domestic pig lineages (rather like human Nia lineages) were replaced during the middle Neolithic with indigenous animals, in little more than five hundred years.
The indigenous pig lineages had spread as far south and east as Armenia by the first millennium BCE.This would not rule out significant acculturation in central Europe during the early Neolithic. Even if the Nia and some of the H lineages were picked up in southeastern Europe, the remaining LBK lineages are quite diverse, including some of potentially Mesolithic western European origin (like Hi). Moreover, simulation studies have suggested that, under realistic demographic parameters for the early Neolithic, the LBK would have had to incorporate a substantial Mesolithic component in order to grow at the archaeologically observed rate.[47] In any event, the virtual lack of Nia (and perhaps also of some of the LBK H lineages) in present-day central European populations suggests that the intrusive LBK populations were subsequently overwhelmed by others, for which the most likely candidates must surely remain their Mesolithic contemporaries, whether local or from elsewhere in Europe. Indeed, direct evidence for late Neolithic assimilation of Mesolithic mtDNA lineages in central Europe has recently been forthcoming.[48] The evidence for the near-extinction of lineages that were common in the early LBK communities is reflected not only in the situation with pig domestication but also in archaeobotanical studies, suggesting that these groups were later replaced.[49] Thus even for central Europe a model of leapfrog pioneer colonization, followed by assimilation of the indigenous hunter-gatherer population, may again be the best explanation for the data. If the replacing populations were intrusive Neolithic populations from elsewhere, their ancestry would in turn be in question.
To clarify the pre-Neolithic picture, we must turn to studies of other parts of Europe, but evidence is thin on the ground (Figure 2.4). Several studies claim to have extracted mtDNA from late Palaeolithic and Mesolithic remains in southwestern Europe.
Here, haplogroup H, which is strikingly absent from pre-Neolithic samples to the northeast, appears in several Iberian Magdalenian samples alongside one with U5. U5b has also been identified in three Iberian Mesolithic samples. Although some of these are dubious, for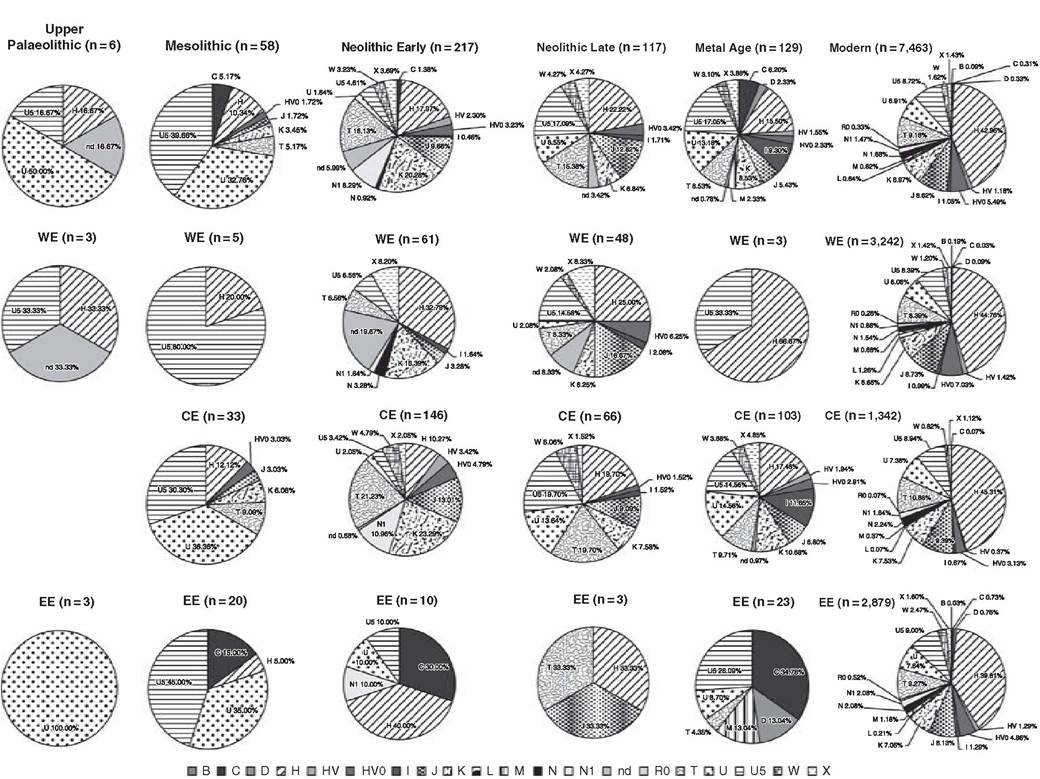
Figure 2.4 Frequencies of mtDNA haplogroups based on ancient DNA from Europe, with comparative contemporary data (April 2014) from the same regions
several reasons, it seems likely that U5 was indeed also present in the southwest before the Neolithic.[50] The presence of H alongside U4 and U5 was also suggested in an earlier study of Portuguese Mesolithic samples,[51] although again we should be cautious of taking these results at face value.
The remainder of the evidence for putative pre-Neolithic lineages comes from northern Europe.[52] Southern Scandinavia only moved into the Neolithic c. 6 ka, and hunter-gatherers such as those of the Pitted Ware Culture (PWC, which actually arose after the arrival of the early Neolithic, and is referred to as middle Neolithic) survived alongside farming groups until c. 4 ka. About a third of nineteen PWC samples carried U4 (previously inferred on the basis of extant variation to have expanded from the eastern European plain after the last glacial maximum[53]), with slightly fewer U5, some T2, a V, and some unidentified lineages. The authors suggest Neolithic or post-Neolithic population replacement in southern Scandinavia, with the PWC arriving by dispersal from the eastern Baltic, with greater continuity from the Mesolithic to the present day in the eastern Baltic region, where U4 in particular occurs at high frequency. Late Mesolithic Russian remains again carry mainly U (U2 as well as U4 and U5), with a single H and some East Asian lineages. The only earlier upper Palaeolithic remains confidently assigned to mtDNA lineages belong to haplogroups U2, pre-U5, pre-U8b, or other U lineages.[54]
There are few data from the early Neolithic of southwestern Europe, and by the late Neolithic at least, as with central Europe, the pattern appears to be close to the modern distribution. N1a has not been found, even in the earliest Neolithic site samples, and the presence of haplogroup U5 suggests at least some assimilation of Mesolithic populations at even the earliest levels. Some intriguing results have emerged from MSY analyses, where there seem to be high levels of lineages that are currently most common in the Near East. These results might suggest a particularly important role for men in the demographic spread of the Neolithic along the Mediterranean. One of these has also been found in the Chalcolithic Tyrolean Iceman, where the high-resolution analysis suggested a possible link to (present-day) Corsica or Sardinia, matching the autosomal evidence.[55]
We have summarized the more reliable available published aDNA data in Figure 2.4. Overall, it does not seem that the results to date can justify claims of a huge genetic discontinuity between the Mesolithic and Neolithic across Europe, or that modern Europeans ‘derive essentially from those Neolithic migrants', as some have claimed. Suggestions that Mesolithic Europe has ‘minimal geographic structure' and that Mesolithic foragers had a ‘common origin' are at best premature, and in part based on the citation of dubious reports in several cases. Although some authors seem to disregard population substructure prior to the Neolithic, this ignores both genetic and archaeological evidence suggesting that the Mesolithic European population was the product of late Glacial and early post-Glacial expansions and subsequent fragmentation.[56] However, the patterns that are emerging from the ancient DNA studies are very striking and intriguing. The implication that European populations began to assume their present genetic structure only from the end of the Neolithic is new and exciting,[57] although Figure 2.4 suggests that it does not yet receive clear support from the data outside of the central Europe LBK zone - rather, the overall picture appears to be one of fluctuations in the frequencies of lineages across both space and time, due in part to genetic drift but very likely also due to sampling.
Recent genome-wide analyses of aDNA - technically a further astonishing development - have been interpreted in a similar way, although presently extrapolated from only half a dozen samples. Three PWC samples from Scandinavia most closely resemble modern Finns and Russians, again (as with the mtDNA) pointing to an eastern source, whereas a single Neolithic Funnel Beaker Culture sample (dating to c. 5 ka, and carrying mtDNA haplogroup H) most resembles southern Europeans. Two Mesolithic samples from Iberia (with identical U5b mtDNAs) appear to fall somewhere between modern central and northern Europeans, and somewhat more distantly from contemporary Iberians.
This might suggest that southern Europe has indeed been reshaped to some extent since late Glacial times by movements from the Near East - perhaps, in part, during the Neolithic - as suggested by mtDNA and Y-chromosome results from modern-day samples - and that the Neolithic involved dispersals from south to north within the continent. Although the major genetic lineages may have been present in Europe since before the Neolithic, their distributions seem likely to have been profoundly reorganized by both long-range and short-range dispersals, by major but regionalized demographic expansions, and by extinctions, such as those of many of the LBK lineages in central Europe. A picture begins to emerge of very diverse histories in different regions, with short-range dispersals (‘leap-frog migrations') and episodes of population coalescence in some cases and replacement in others, fitting radiocarbon evidence for short bursts and pauses in the process of Neolithic spread.
Ancient DNA is starting to contribute to the study of other aspects of the spread of farming. For example, present-day Europeans display high levels of lactose persistence (the ability of adults to digest milk sugar) due to the presence of a particular DNA variant, which differs from the variant conferring persistence where it occurs in other parts of the world and most likely rose to high frequencies due to selection in early European dairying populations. One Mesolithic and eight early Neolithic samples from central and northeastern Europe all lacked the persistence variant, supporting the view that dairying had to become embedded in the culture before the variant could be selected.[58] It is likely that this is only the beginning of a dissection of the impact of the spread of farming on the human genome, and vice versa.