85 Malaria
Malaria is the disease resulting from infection by one or more of four species of protozoan parasites of the genus Plasmodium. These parasites are normally transmitted from one human host to the next by the bite of an infected female mosquito of the genus Anopheles.
Although malaria has receded from many temperate regions in this century, the disease continues to be a major cause of morbidity and mortality in many tropical and subtropical countries. Three of the species — Plasmodium υiuax, Plasmodium falciparum, and Plasmodium malariae - are widely distributed; the fourth, Plasmodium oυale, is principally a parasite of tropical Africa. P. υiυax (the agent of benign tertian malaria) and P. falciparum (causing malignant tertian malaria) are responsible for the great majority of cases and deaths attributed to malaria throughout the world.Malaria is characteristically paroxysmal, and often periodic. The classical clinical episode begins with chills, extends through a bout of fever, and ends with sweating, subsiding fever, a sense of relief, and, often, sleep. Between the early paroxysms the infected person may feel quite well; as the disease progresses, however, the patient may be increasingly burdened by symptoms, even in the periods between febrile paroxysms. Although infection by any species may have serious, even fatal, consequences, P. falciparum infection is particularly dangerous because of complications associated with this parasite.
The term malaria, from the Italian mala and aria (“bad air”), was certainly in use in Italy by the seventeenth century to refer to the cause of intermittent fevers thought to result from exposure to marsh air or miasma. Horace Walpole wrote home from Italy in 1740 about “[a] horrid thing called mal’aria, that comes to Rome every summer and kills one.” This is said to be the first use of the term in English, and it may well have been the first appearance of the word in print (Russell et al.
1963). However, Walpole and other writers of the eighteenth century, and later, used the term to refer to the presumed cause rather than the disease. It was only after the pathogenic agents were identified at the end of the nineteenth century that usage shifted so that the term “malaria” came to refer to the disease rather than the agent.Etiology
Protozoa assigned to the genus Plasmodium are parasitic in many species of vertebrate animals including birds, reptiles, amphibians, and mammals. Most Old and New World primate species serve as hosts for plasmodia, and some of these parasites are closely related to the species established in human populations. It is generally accepted that the human malaria parasites evolved in association with early human beings, perhaps differentiating into the four species recognized today in the mid-Pleistocene (Gamham 1966; Bruce-Chwatt and Zulueta 1980).
All of the mammalian plasmodia have similar two-phase life cycles: an asexual (schizogonic) phase in the vertebrate host and a sexual (sporogonic) phase in female Anopheles mosquitoes. These cycles reflect ancient anopheline and vertebrate hostparasite relationships that seem to date from at least the Oligocene.
The sexual phase in the mosquito is initiated as the female anopheline takes a blood meal from a human or other vertebrate host. Parasites in ingested red blood cells are released as male and female gametes in the stomach of the mosquito. Fusion of the gametes produces a zygote, which, after rapid development, migrates and encysts in the stomach wall. At maturity, after some 10 to 20 days, this oocyst releases thousands of sporozoites, motile forms that migrate to the salivary glands. When the now infective mosquito bites and probes for vertebrate blood, many of these sporozoites are likely to be injected into the new host in released saliva. The duration of the sexual or Sporogonic phase of the cycle is controlled principally by environmental temperature and humidity.
P. vivax requires a minimum temperature of 16oC; P. falciparum, at least 18oC. The duration of P. vivax sporogony is 9 to 16 days and that of P. falciparum, 12 to 24 days, when environmental temperatures range between 20o and 25oC (Bruce-Chwatt 1987).Most of the sporozoites injected by the biting mosquito are phagocytosed, but some, traveling in the bloodstream, reach the liver, enter parenchymal cells, and (as tissue schizonts) proceed by nuclear division to form large numbers (10,000 to 20,000) of merozoites. Eventually, about 6 to 16 days after initial infection, the preerythrocytic schizont ruptures, releasing its merozoites into the bloodstream. Those merozoites that escape phagocytosis invade red blood cells and, as trophozoites, initiate the erythrocytic portion of the asexual phase. The trophozoite form breaks down the hemoglobin of the host erythrocyte, leaving hematin pigment as a digestive product. As it grows, the parasite (now called an erythrocytic schizont) divides, producing 8 to 24 merozoites, depending on the species. For the human malaria parasites, schizogony in the red cell lasts about 48 hours or, for P. malariae, about 72 hours. When the schizont reaches maturity, the erythrocyte bursts and the merozoites are released. Again some will be phagocytosed whereas others will invade uninfected red cells. Erythrocytic schizogony may then continue through repeated cycles with increasing synchronism, manifest as clinical periodicity. As a consequence, more and more red cells are parasitized and destroyed. Immune responses or therapeutic intervention can check the process short of profound anemia, complications, and death.
Following clinical recovery, P. υiυax and its close relative P. ovale are capable of causing relapse, even after several years. Recrudescence of quartan malaria (P. malariae infection) may occur after many years, even 30 or more years after initial infection. P υiυax and P ovale infections can persist because of the survival in the host liver of dormant stages capable of reinitiating erythrocytic schizogony.
Recrudescent P malariae infection appears to be due to longterm survival of erythrocytic-stage parasites.P. falciparum parasites, on the other hand, are much more limited in time of survival; if the disease is left untreated and is not fatal, P. falciparum infection will terminate spontaneously without relapse, usually in less than a year. Instead of developing as trophozoites, small numbers of red cell merozoites differentiate as male and female forms (gametocytes). These circulate in erythrocytes and may eventually be ingested by a female Anopheles mosquito - to begin a new sexual phase of the life cycle.
Although malaria transmission normally follows the bite of the infected mosquito, several other routes have been recorded, including congenital transmission and direct transfer of parasites by blood transfusion or by contaminated needles shared by drug abusers. Before the advent of antibiotic therapy, malaria infections were also sometimes induced by injection for therapeutic purposes - for example, to produce high fevers in the treatment of late-stage syphilis.
Distribution and Incidence
Although the eradication campaigns of the 1950s and 1960s contributed to substantial declines in the incidence of malaria in many countries, and to the elimination of transmission in some, malaria persists as a major contributor to ill health in vast areas of the tropical and subtropical world. Reported cases of malaria are increasing from year to year, especially in areas of Asia and the Americas undergoing agricultural colonization with forest clearing and pioneering of unexploited lands. Eradication campaigns have given way to long-term control programs in most areas where the disease remains endemic, and in some countries control is now being linked to or integrated with systems of primary healthcare (Bruce-Chwatt 1987; Hilton 1987).
The present global situation has been categorized by David Clyde (1987) as follows:
1. Areas where malaria never existed or disappeared spontaneously (with a current population totaling some 1.3 billion)
2.
Areas where control and eradication campaigns, in combination with other factors, have eliminated endemic malaria in recent years (population about 800 million)3. Areas under continuing control, including most endemic countries of Asia and the Americas (population about 2.2 billion)
4. Areas with little or no organized control, mainly in Africa south of the Sahara (population about 400 million)
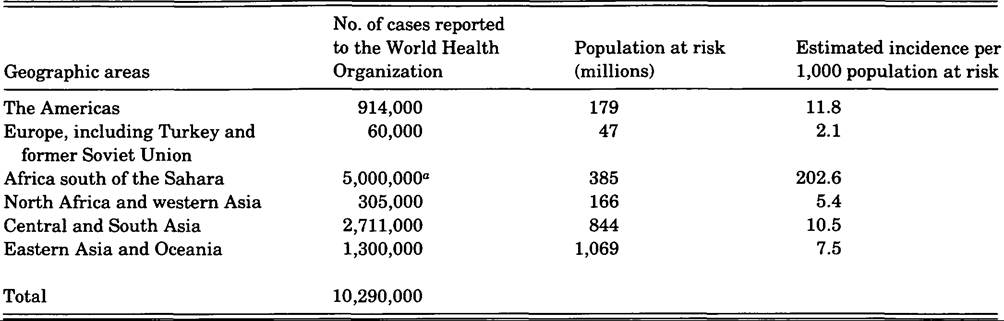
“Estimated; mainly clinical diagnoses.
Source: Adapted from World Health Organization data and Clyde (1987, tables 1 and 2).
Reported cases, and estimates for Africa south of the Sahara, are summarized in Table VIII.85.1 for 1984, together with incidence estimates. The total of some 10 million cases probably represents less than 10 percent of actual cases for that year because of reporting and surveillance deficiencies. Clyde (1987) believes that the infected population in 1984 may actually have exceeded 100 million in a population at risk of 2.6 billion. Malaria-related deaths are also poorly reported; David Hilton (1987) notes that worldwide estimates of deaths are still in the range of hundreds of thousands every year.
The Americas. The United States, Canada, and most countries of the Caribbean are essentially free of malaria transmission and report only small numbers of imported cases. Costa Rica, Panama, and several southern South American countries are also nearly free of local transmission. Haiti and the Dominican Republic report only P. falciparum cases, although in substantial numbers. P. vivax is the prevailing species in Mexico, Central America, and northern South America, although many P. falciparum cases are also reported, especially from Brazil, Colombia, and Ecuador.
Europe, Turkey, and the former Soviet Union. Malaria is no longer endemic in Europe and the Soviet Union, but Turkey still reports some autochthonous cases.
Africa south of the Sahara.
About 90 percent of the population in this region is still at risk, and transmission is high in many areas, especially in rural West Africa (Spencer 1986). P. falciparum is the predominant species.North Africa and western Asia. Only a few small foci of autochthonous transmission persist in this region.
Central and South Asia. Malaria remains endemic in most countries of the region. P. vivax is predominant, but P. falciparum is also important, and this species appears to be increasing in relative prevalence (Spencer 1986).
Eastern Asia and Oceania. The northeastern area of Asia, including Japan, is free of transmission, as are many of the smaller islands and island groups in Oceania. Endemic foci persist in some of the larger island areas (e.g., the Philippines, the Solomon Islands, Papua New Guinea, and Indonesia) and in most of mainland Southeast Asia and China. P. υiυax prevails in China, where the incidence is steadily declining. In Thailand, which is also experiencing a decline in incidence, P. falciparum is somewhat more prevalent than P. vivax.
Epidemiology and Control
Malaria transmission in any locale depends upon the complex interactions of parasites; vector mosquitoes; physical, socioeconomic, and environmental factors; and human biology, demography, and behavior.
The four species of plasmodia differ in many biological characteristics. Each species, for example, has somewhat different environmental temperature requirements for sporogony; each is different in many features of its Schizogonia cycle, and as a consequence, each has distinctive clinical manifestations. P. falciparum differs so much from the other three species (of the subgenus Plasmodium) that it has been assigned to its own subgenus, Laverania (Gamham 1966). Within each species, variation in strains is also important. Differences in strains influence sporogonic development in mosquitoes, virulence and thus the clinical course, and the development of resistance to antimalarial drugs.
The presence of anopheline mosquitoes capable of supporting sporogony is fundamental for natural malaria transmission. Although more than 400 species of Anopheles have been described, natural sporozoite infections have been found in only about 67 species, and only some 30 species are recognized as important vectors (Spencer 1986). Usually, in any endemic area, only one, two, or three vector species are responsible for most of the malaria transmission. Parasite species and strain differences affect anopheline receptivity to infection. Vector competence for transmission is also influenced by mosquito population densities, flight ranges, feeding and resting habits, blood meal preferences (anthropophilic for human blood; zoophilic for animal blood), blood meal frequencies, and the lifespan of the female anopheline. In recent decades, resistance to insecticides has also become a major vector-related determinant of transmission.
Vector, parasite, and host interactions are profoundly affected by factors in the environment. Physical factors, such as temperature, rainfall, and humidity, control mosquito survival and the duration of sporogony. Other factors such as latitude, altitude, and landscape characteristics (natural or human-modified) influence mosquito breeding, vector densities, and vector survival, as well as many kinds of human behavior. In addition, features of the social and economic environment contribute in many ways to patterns of transmission.
Age, gender, and a variety of human demographic variables, together with genetic factors, immune mechanisms, and health conditions (e.g., nutritional status, pregnancy, and concurrent infections), constitute the final host-related set of determinants in the malaria epidemiological complex that must be well understood before fully effective control can be instituted in any setting.
The incidence and prevalence of clinical cases may be useful measures for describing the status of malaria in an area, but parasite and spleen rates are even more helpful for determining the level of endemicity - spleen rates particularly, because enlarged spleens are indicative of past or recent active erythrocytic schizogony. The spleen rate is the proportion of persons, usually children 2 to 9 years old, with enlarged spleens at a particular time, whereas the parasite rate is the proportion of persons in a population with microscopically confirmed parasitemia at a particular time.
Malaria endemicity is classified on the basis of spleen or parasite rates:
1. Hypoendemic malaria: spleen or parasite rates up to 10 percent in 2- to 9-year-old children
2. Mesoendemic malaria: spleen or parasite rates of 11 to 50 percent in 2- to 9-year-olds
3. Hyperendemic malaria: spleen or parasite rates over 50 percent in 2- to 9-year-olds together with a high adult spleen rate
4. Holoendemic malaria: spleen or parasite rates over 75 percent in 2- to 9-year-olds with low adult spleen rates and high infant parasite rates
In a pioneering work, George Macdonald (1957) attempted to fit many of the variables noted above into an epidemiological model. His ideas continue to be influential in contemporary malariology despite many advances in epidemiological and mathematical modeling sophistication. Macdonald’s principles in defining stable and unstable malaria, for example, continue to be helpful in studies of epidemiological patterns. Stable malaria is characteristically endemic, often hyper- or holoendemic. There is little seasonal change; transmission continues through most or all of the year; epidemics of malaria are very unlikely to occur; and most of the population, except infants, has some immunity as a result of experience with the disease. Control under these circumstances is likely to be very difficult. Malaria in much of tropical Africa south of the Sahara corresponds to Macdonald’s stable extreme.
Unstable malaria may be endemic, but transmission varies from year to year and may be strictly seasonal, reflecting the strong seasonal change typical in unstable areas. Seasonal epidemics may occur, collective immunity varies and may be low, and children as well as infants may be nonimmune. Control is relatively easy, and indeed, many of the coιmtries only recently freed from endemic malaria were in the unstable category.
Immunity
Acquired and innate immunity are important factors in the epidemiology of malaria. Innate resistance has been recognized in some human populations. In parts of Central and West Africa, many individuals lack the Duffy blood group antigens Fya and Fyb and are thus resistant to P. υiυax infection. Generally, in these areas, P. ovale replaces P. υiυax as the prevailing cause of benign tertian malaria. In endemic P. falciparum areas of Africa, those individuals heterozygous for hemoglobin AS or sickle trait are more likely to survive malignant tertian ma- Iaria than are hemoglobin A homozygotes. The persistence of sickle-cell disease (in hemoglobin SS homozygotes), with its unfortunate consequences, is balanced by the substantial advantage conferred by the AS condition, but only as long as P. falciparum remains endemic in the area. Several other hemoglobin variants have been thought to provide some protection against P. falciparum infection; it is known that persistent hemoglobin F (fetal Hb) delays development of falciparum parasites (Bruce-Chwatt 1987). Another genetic condition, deficiency of the red blood cell enzyme glucose 6-phosphate dehydrogenase (G6PD), also provides a degree of protection against falciparum infection.
Acquired immunity in malaria is species-specific and also specific to stage, that is, to the sporozoite, to the asexual forms in the blood, or to the sexual stages (Clyde 1987). In endemic areas, newborns may be protected for a few months by maternal antibodies that have crossed the placenta. After this phase of protection, however, infants and toddlers are especially vulnerable; most of the severe illnesses and deaths due to malaria in endemic regions occur in these early years. Older children and adults gradually acquire immunity with repeated exposure to infection. This partial immunity (premunition) is sustained in the presence of low densities of parasites in the blood. In premune individuals the spleen may remain enlarged; hence the spleen rate may be relatively high in an adult population despite a low parasite rate. With prolonged exposure and infection, however, as in a holoendemic area, eventual spleen scarring and shrinkage leads to a low adult spleen rate. Attenuated malaria certainly contributes to impaired resistance to other diseases; thus improved malaria control in endemic areas generally results in declines in morbidity and mortality from all causes (Bruce-Chwatt 1987).
Clinical Manifestations and Pathology The classical malaria paroxysm is initiated by a chill, lasting up to an hour and often accompanied by headache, nausea, and vomiting. The chill is followed by a period of spiking fever lasting several hours. The subsiding fever is accompanied by sweating, often profuse. The relieved patient may drift off to sleep, to awaken feeling relatively well. The early paroxysms may be asynchronous.' As the clinical episode progresses, P. malariae paroxysms occur about every 72 hours (quartan periodicity), whereas those of the other three species occur with tertian (48hour) periodicity. P. falciparum infection is less likely to become clearly synchronous, and the classic clinical picture may never develop. Mixed infections of two or even three species may produce atypical and confusing clinical manifestations.
These clinical events are related to the behavior of the parasites in the bloodstream. The onset of chill reflects the escape of parasites and metabolic products from great numbers of ruptured red cells. By the end of the paroxysm, the parasites will have invaded new red cells to initiate another generation of erythrocytic schizogony. With each cycle of invasion, multiplication, and rupture, more red cells are destroyed. In the absence of treatment and immune defenses, severe anemia may develop, particularly in falciparum malaria. The spleen enlarges during the acute episode, in part because of its function in filtering out of the bloodstream the detritus resulting from red cell destruction. The liver may also increase in size.
Although malaria caused by the other three species may be fatal, most of the serious complications and deaths are caused by P. falciparum. This species can invade a high proportion of the total red cell mass, and the parasitized cells tend to agglutinate, occluding capillaries in almost any organ. These occlusions are considered to underlie cerebral, gastrointestinal, adrenal, and other complications in falciparum malaria. Cerebral malaria begins with headache and may progress to convulsions, delirium, and coma. Gastrointestinal complications resemble bacillary dysentery or cholera.
The initial uncomplicated and untreated episode of malaria usually lasts 2 to 4 weeks (about twice as long for P. malariae). Relapses commonly occur in untreated malaria. Drug-resistant strains of P. falciparum may also cause relapses after inadequate treatment.
Blackwater fever is a distinctive intravascular hemolytic disorder associated with longstanding and repeated falciparum malaria infection, especially in hyper- and holoendemic areas. The incidence of blackwater fever has fallen considerably in the second half of this century as malaria control has improved and as drug prophylaxis and treatment has expanded from an earlier dependence on quinine. Quinine is not an essential etiologic factor, but the onset of blackwater fever has often been noted to follow the administration of therapeutic doses of the drug. It is also known that the disorder rarely occurs in persons who have taken quinine regularly as a prophylactic. Blackwater fever is characterized by abrupt onset, chill, fever, jaundice, nausea, vomiting, and red or dark-brown urine containing hemoglobin. The case mortality rate, resulting principally from anuria and uremia, usually falls between 10 and 30 percent.
History and Geography
Since at least the mid-Pleistocene, many thousands of generations of humans have been parasitized by the plasmodia (Garnham 1966). Species very similar to the human plasmodia have also been described from gorillas and chimpanzees; indeed the relationships of the malaria parasites of humankind and the African apes are extremely close (Dunn 1966; Gamham 1966). It is therefore certain that human malaria is an Old World disease in its origins. There has, however, been some debate about the timing of malaria’s appearance in the Western Hemisphere (McNeill 1976). Some have suggested that malaria was present in the New World long before the time of European contact, but a strong case can be made that the hemisphere was malaria-free until the end of the fifteenth century (Dunn 1965; McNeill 1976).
Malaria could have reached the New World before 1492 only as an infection of migrants from northeast Asia or by pre-Columbian sea-borne introductions. The possibility that humans brought malaria overland into North America from Siberia can almost certainly be discounted; conditions for malaria transmission were unsuitable in the far north during and after the Pleistocene, as they are today. It is equally unlikely that the Vikings could have introduced malaria in the centuries before Columbus. These voyagers came from regions of Europe and the North Atlantic (Greenland, Iceland) presumably free of malaria at that time, and they seem to have visited northeastern North American coasts that were north of any possible receptive anopheline mosquito populations. Similarly, any voyagers landing on American shores from the central or eastern Pacific could not have carried the parasites with them because islands in that region are free of anopheline vectors and thus of locally transmitted malaria. Voyagers reaching American coasts from eastern Asia (e.g., fishermen adrift from Japan) could conceivably have introduced malaria, but this possibility too is remote.
Moreover, and more to the point, colonial records indicate that malaria was almost certainly unknown to the indigenous peoples of the Americas. It is also evident that some areas that supported large preColumbian populations soon became dangerously malarious after European contact (McNeill 1976). The absence in aboriginal American populations of any of the blood genetic polymorphisms associated with malaria elsewhere in the world is another kind of evidence consistent with the conclusion that the Western Hemisphere remained free of the disease until contact (Dunn 1965).
In the first two centuries after 1492, malaria parasites must have been introduced many times from Europe and Africa. Anopheline vectors were at hand, species native to the Americas. Together with smallpox, measles, and other infectious diseases brought across the Atlantic from the Old World, malaria soon began to contribute to the depopulation of the indigenous peoples, especially those of the lowland American tropics (Borah and Cook 1963; Crosby 1973; McNeill 1976).
From its early, usually coastal, sites of introduction, malaria spread widely in North, Central, and South America, limited principally by altitude and latitude - that is, by factors controlling the distribution of vector mosquitoes. By the nineteenth century in North America, the disease was prevalent in much of the Mississippi Valley; seasonal transmission occurred even in the northernmost areas of the Mississippi basin (Ackerknecht 1945). Malaria transmission, unstable and seasonal, also extended into the northeastern United States, well north in California, and far to the south in South America. By the eighteenth and nineteenth centuries, malaria had also become established as a stable endemic disease in the American subtropics and tropics, including most of the islands of the Caribbean.
The Old World gave malaria to the New; the New World, however, provided the first effective remedy for the disease. It is recorded that a sample of cinchona bark, taken from a Peruvian tree as a medicinal, was carried to Europe in 1632 by a Spanish priest (Russell et al. 1963). The bark was soon discovered to provide relief from certain intermittent fevers. This therapeutic action allowed Richard Morton and Thomas Sydenham in England in 1666, and Francesco Torti in Italy in 1712, to begin the process of defining malaria as a clinical entity separable from other fevers, which failed to respond to cinchona (Russell et al. 1963). By the end of the seventeenth century, cinchona bark was an important export product from Peru, well established in the treatment of intermittent fevers.
The human malaria parasites have evolved in association with their evolving hosts and have followed the human species through most, but not all, of its dispersal in Africa, Asia, and Europe. Transmission, dependent on suitable anopheline mosquitoes, has always been limited by all those environmental conditions, influenced by latitude and altitude, that control vector breeding and survival.
The antiquity of the human—Plasmodium association in the Old World is symbolized, biologically, by the existence in modern populations of some of the blood genetic polymorphisms. Early texts - Chinese, Hindu, Chaldean, Greek - give us other evidence of this ancient association (Russell et al. 1963; McNeill 1976; Bruce-Chwatt and Zulueta 1980). Malaria was probably endemic in Greece by the fourth century, B.C.; Hippocrates described the types of periodicity of intermittent fevers - quotidian, tertian, quartan - and took note of the enlargement of spleens in those who lived in low, marshy districts. In Italy, too, the intermittent fevers were well known, for example, to Cicero, and well described by Celsus, Pliny the Elder, and Galen (Russell et al. 1963).
Malaria was certainly a disease of some importance in the centuries of Roman domination of Europe and the Mediterranean basin. However, in their review of malaria in early classical times, especially in relation to military operations, L. J. Bruce- Chwatt and J. de Zulueta (1980) reached the conclusion that malaria was much less destructive than it has been in recent centuries, primarily because P. falciparum was absent or rare and the other species were less intensely transmitted. The conclude that P. falciparum failed to spread in those centuries because Anopheles atroparυus was refractory as a vector for the falciparum strains then introduced from time to time by travelers. A. atroparυus, basically a Zoophilic species, was also a poor vector for P. malariae and P. υiυax. By late classical times, however, it appears that two other anopheline species, Anopheles Iabranchiae and Anopheles sacharoυi, had been introduced and dispersed along the coasts of southern Europe from their North African and Asian origins. These anthropophilic species were much more effective as vectors for all three of the species of plasmodia. By the final centuries of the Roman Empire, malaria was a more lethal force and may have contributed to the social, political, and cultural decline that had set in.
After the fall of the Roman Empire, the history of malaria in Europe and the Mediterranean region is obscure for many centuries (Bruce-Chwatt and Zulueta 1980). With few exceptions medieval medical writers provide only sketchy and confusing accounts of cases and outbreaks that may have been due to malaria. During the Renaissance, the historical record is clearer and richer, but malaria appears not to have represented a major problem to society in those centuries. It is not until the seventeenth and eighteenth centuries that malaria became resurgent in Europe, not only in the south but, in periodic outbreaks, as far north as the Netherlands, Germany, southern Scandinavia, Poland, and Russia (Bruce-Chwatt and Zulueta 1980).
Through all of these centuries the record of malaria’s human impact in Asia and Africa is fragmentary. With the onset of European colonization, however, it soon became obvious that endemic malaria was a threat almost everywhere in the Old World tropics, especially to the colonizers (who were, of course, generally nonimmune upon arrival in the tropics). The beginnings of tropical medicine and modem malariology are entangled in this recognition (Dunn 1984).
The modem era in malariology began in the last two decades of the nineteenth century with the identification of the causal parasites and the recognition of the role of anopheline mosquitoes as vectors. These discoveries provided the rationale for new strategies in malaria control, developed during the first third of the twentieth century. Malaria control itself was not a new concept; nor was the control of mosquitoes a new idea at this time. Humankind had sought from ancient times to control biting mosquitoes as pests (Russell et al. 1963). The new rationale, however, provided for much more specific vector control, directed principally at those anophelines that proved to be important in transmission. Malaria control was further strengthened in the 1930s with the introduction of synthetic antimalarials, useful not only in treatment but also in prophylaxis. The 1940s brought further advances in chemotherapy together with the first of the residual insecticides, DDT.
By the late 1940s and early 1950s, the resources for treatment and control of malaria appeared to be sufficiently formidable to justify attempts at national eradication in a few countries (e.g., Venezuela, Italy, the United States). Early success in local or aerial eradication, based on residual spraying, together with concerns about the emergence of anopheline resistance to DDT and other insecticides, led to a decision in 1955 to commit the World Health Organization to malaria eradication. This commitment pushed many countries into supposedly timelimited eradication campaigns, often with considerable WHO financial and adivsory support. Some of these campaigns were successful, especially in countries with unstable malaria, but others faltered after dramatic initial reductions in incidence. By the end of the eradication era, in the early 1970s, some hundreds of millions of people were living in areas where campaigns had eliminated endemic malaria (Clyde 1987), but in many other areas malaria continued to prevail, forcing a return to long-term control strategies.
In 1980 the World Health Organization began to recommend that malaria control be coordinated with primary health care. The experiences, especially the difficulties, of the eradication years also made it imperative that modern malaria control be strongly supported by epidemiological field work with recognition of local variability in transmission patterns and of sociocultural, economic, and human behavioral influences on transmission and control (Bruce-Chwatt 1987). Malaria control and therapy continues to be complicated by mosquito resistance to insecticides and by parasite resistance to drugs. Vaccine development proceeds, although slowly. The problems posed by malaria persist, but they are not insoluble. Malaria will remain endemic in many countries in the twenty-first century, but the prospects are good for steady improvement in control.
Frederick L. Dunn
Bibliography
Ackerknecht, E. H. 1945. Malaria in the Upper Mississippi Valley, 1860—1900. Bulletin of the History of Medicine, Supplement No. 4.
Borah, Woodrow W., and Sherburne F. Cook. 1963. The aboriginal population of Central Mexico on the eve of the Spanish Conquest. Ibero-Americana No. 45.
Bruce-Chwatt, L. J. 1987. Malaria and its control: Present situation and future prospects. Annual Review of Public Health 8: 75-110.
Bruce-Chwatt, L. J., and J. de Zulueta. 1980. The rise and fall of malaria in Europe: A historico-epidemiological study. London.
Clyde, David F. 1987. Recent trends in the epidemiology and control of malaria. Epidemiologic Reviews 9: 219-43.
Crosby, Alfred W., Jr. 1973. The Columbian exchange: Biological and cultural consequences of 1492. Westport, Conn.
Dunn, Frederick L. 1965. On the antiquity of malaria in the Western Hemisphere. HumanBiology 37: 385-93.
1966. Patterns of parasitism in primates: Phylogenetic and ecological interpretations, with particular reference to the Hominoidea. Folia Primatologica 4: 32945.
1984. Social determinants in tropical disease. In Tropical and geographical medicine, ed. K. S. Warren and A. A. F. Mahmoud, 1086—96. New York.
Garnham, P. C. C. 1966. Malaria parasites and other haemosporidia. Oxford.
Hilton, David. 1987. Malaria: A new battle plan. Contact 95: 1-5.
Macdonald, George. 1957. The epidemiology and control of malaria. London.
McNeill, William H. 1976. Plagues and peoples. Garden City, N.Y.
Russell, P. F., et al. 1963. Practical malariology, 2d edition. London.
Spencer, Harrison C. 1986. Epidemiology of malaria. In Clinics in tropical medicine and communicable diseases, Vol. 1: Malaria, 1—28.